Семейство отличается от других групп афиллофоровых грибов отсутствием пряжек на гифах, постоянным присутствием бурых пигментов, наличием щетинок и щетинкоподобных элементов в гимении и ткани, большим по сравнению с другими группами количеством видов, патогенных для живых деревьев. Гименохетовые грибы характеризуются тенденцией к паразитизму и к относительно узкой специализации по породам-хозяевам.
По макроморфологическому строению базидиомы гименохетовых грибов относятся ко всем известным морфологическим типам: резупинатному, распростерто отогнутому, сидячему и дифференцированному на шляпку и ножку. Распростертые базидиомы являются истинно резупинатными, или распростерто отогнутые и сидячие при горизонтальном положении субстрата могут образовывать распростертые формы. У истинно резупинатных базидиом край тонкий, сходящий на нет, или толстый, валиковидный, но, как правило, плотно приросший. Ризоморфы по краю наблюдаются только у одного гриба из рода Inonotopsis. Подстилка у резупинатных гименохетовых грибов очень тонкая до почти отсутствующей. Трубочки на вертикальном или наклонном субстрате иногда скошенные. Резупинатные базидиомы могут прикрепляться к субстрату в нескольких точках или плотно к нему прирастать.
Грибы с распростерто отогнутыми базидиомами морфологически наиболее пластичны. Виды, входящие в эту группу, способны давать все конфигурации - от полностью распростертых базидиом до сидячих, с широким или суженным (ножковидным) основанием. Распростерто отогнутая форма образуется путем отгиба верхнего края тонкой, раковинообразной или толстой, желвакообразной, копытовидной или консолевидной базидиомы. В качестве видового признака может фигурировать большая или меньшая массивность базидиом. Что же касается их геометрической формы, то описать ее почти невозможно, так как вертикальный, скошенный или ущемленный субстрат способен стимулировать образование самых причудливых конфигураций. У базидиом этой группы гименофор, особенно многослойный, более или менее стабилизирован, т.е. они почти не образуют скошенных трубочек. Положительный геотропизм гименофора достигается за счет неравномерного роста бесплодной ткани базидиомы. По количеству относящихся сюда видов эта группа самая многочисленная. Примерами могут служить виды родов Hymenochaete, Phylloporia, Inonotus, Phellinus с однослойным и слоистым гименофором.
Сидячие копытовидные и консолевидные базидиомы без ножек, как правило с широким основанием, развиваются на живых стволах или на вертикально стоящем сухостое или пнях. Сюда относятся как однолетние, так и многолетние виды. Типичные представители этой группы имеют базидиомы с массивной хорошо развитой трамой (Inonotus hispidus, Phellinus igniarius).
Вид Onnia triqueter благодаря суженному основанию представляет собой переход к последней группе, в которую входят грибы с базидиомами, дифференцированными на шляпку и ножку. Эта морфологическая группа объединяет виды различного происхождения, развивающиеся как на почве, так и на древесине. При росте на почве, корнях деревьев и горизонтальном валеже они образуют боковую, центральную или эксцентрическую ножку. Изгиб боковой ножки при наличии скошенного или вертикального субстрата обеспечивает правильную, положительно геотропичную ориентацию гименофора, без изменения формы шляпки. К этой группе относятся представители родов Onnia, Coltricia. Вид С. perennis часто образует шляпки, срастающиеся краями.
Деление это условное так как между соседними группами имеются переходы. Виды, характеризующиеся распростерто отогнутыми базидиомами, могут образовывать и совершенно распростертые, и сидячие формы (Inonotus radiatus, Phellinus igniarius, Ph. conchatus и др.). У грибов рода Onnia имеются переходы между сидячей формой и дифференциацией на шляпку и ножку. Для каждого таксона форма базидиомы варьирует в пределах, ограниченных его наследственностью.
Консистенция ткани обусловлена типом гифальной системы, количественным соотношением гиф разных типов и характером их расположения в базидиоме. У гименохетовых грибов известны мономитический и димитический типы гифальных систем. Плотное параллельное расположение скелетных толстостенных гиф обусловливает деревянистую консистенцию базидиом Phellinus igniarius, несколько более рыхлое, переплетающееся расположение — пробково-деревянистую консистенцию Ph. conchatus, преобладание более тонкостенных генеративных гиф — пробковые базидиомы Ph. gilvus. Мономитическое, с относительно плотным расположением гиф строение ткани обеспечивает кожистую консистенцию видов рода Hymenochaete. Представители рода Inonotus характеризуются пучковым расположением гиф: генеративные гифы с утолщенными стенками располагаются параллельно в переплетающихся тяжах, образуя подобие сети, в ячейках которой рыхло переплетаются одиночные тонкостенные гифы (I. hispidus, I. pseudohispidus и др.). Такое строение обеспечивает возможность накопления запасов влаги. Плотное параллельное расположение относительно тонкостенных гиф мономитического рода Coltricia дает кожистую консистенцию тонких шляпок этого гриба.
Окраска ткани служит важным диагностическим признаком у трутовиков. Представители семейства Hymenochaetaceae имеют темно-бурую, рыже-бурую или бурую окраску базидиом. Это хороший диагностический признак семейства, но в пределах родов, особенно крупных, встречаются все оттенки указанных окрасок.
Представители семейства гименохетовых характеризуются гладким, шиловидным, трубчатым, лабиринтовидным и концентрически пластинчатым гименофором. Гладкий гимений имеют представители рода Hymenochaete, шиповидный — родов Astero-don и Hydnochaete. Концентрически пластинчатый, неслоистый гименофор отличает тропический род Cyclomyces. Грибы остальных родов семейства характеризуются трубчатым гименофором. Базидиомы большинства видов рода Phellinus способны к многолетнему существованию с образованием слоистого гименофора. Иногда трубочки отделяются друг от друга прослойками стерильной ткани (Ph. hartigii). Ряд видов этого рода существует в течение 1—3 сезонов и имеет однослойный гименофор. Представители родов Phylloporia, Coltricia, Onnia, Inonotus однолетние, гименофор всегда однослойный.
Поры у грибов с трубчатым гименофором, иногда лабиринто-видные (Phellinus pini) или неправильные (Phylloporia ampelina), чаще округлые или угловатые, цельнокрайные или с зубчато рассеченными краями, в молодом возрасте часто опушенными. Рассеченные зубчатые края пор наиболее характерны для рода Inonotus. У некоторых видов распростертые базидиомы регулярно образуются на вертикальном субстрате; трубочки в этом случае скошенные (I. obliquus).
Слоистость и конфигурация гименофора служат признаками родового и видового ранга. Средняя величина пор и количество их на линейный миллиметр поверхности гименофора у трубчатых родов относятся к числу стабильных видовых признаков и играют существенную роль в определении. Для умеренной и холодной климатических зон характерны поры средней величины (3— 6 на 1 мм), для тропиков или очень крупные (1—2 мм в диам.), или, напротив, очень мелкие (8—12 на 1 мм).
Верхняя поверхность шляпок, ее строение и окраска также характерны для каждого вида. У гименохетовых грибов плотная корка на поверхности отсутствует, хотя поверхностный слой обычно бывает уплотнен или, наоборот, более мягкий, войлочный по сравнению с прилегающим слоем ткани. Голая (без корки и кожицы) поверхность шляпок гименохетовых грибов бывает гладкой, шероховатой, бархатистой, войлочной, волосистой, грубо-щетинистой (I. hispidus), ровной или концентрически-бороздчатой (Phellinus conchatus), что связано с неравномерным ростом базидиомы.
Микроморфологическое строение базидиом определяется типом гифальной системы, характером гиф и их расположением.
Гифы поверхности шляпки и ножки часто отличаются по расположению, толщине стенок, длине клеток от гиф ткани. Как правило, клетки этих гиф более короткие, гифы опушения склеены в плотные пучки, гифы голой гладкой или шероховатой поверхности часто агглютинированы.
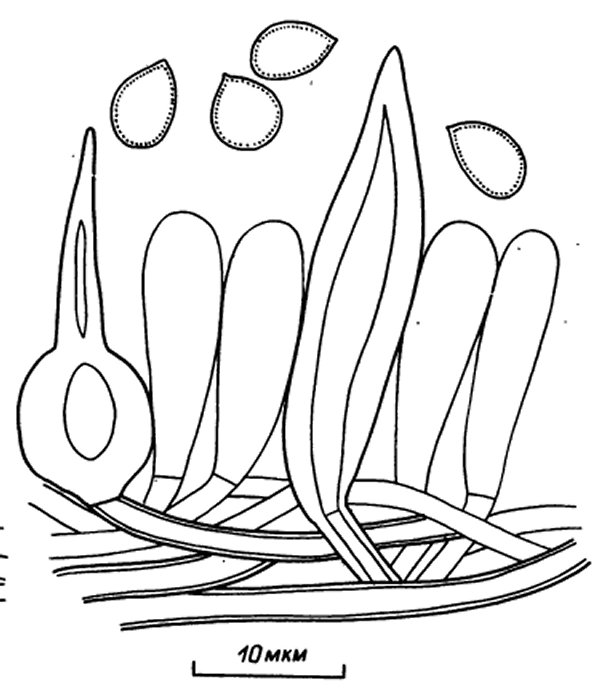
Гифы ткани и гименофора, согласно концепции Корнера (Corner, 1948), дифференцированы на типы, которые он назвал скелетными, связывающими и генеративными. Генеративные гифы, как показывает само название, развиваются первыми и служат исходным материалом для дифференцировки других типов гиф и гимения. Это гифы неограниченного роста, развивающиеся в зоне роста, тонкостенные, с пряжками или анастомозирующими гифами, обычно сильно ветвящиеся. У некоторых групп, в том числе и семейства гименохетовых, пряжки на генеративных гифах отсутствуют. Иногда генеративные гифы имеют вторично утолщенные стенки и в этом случае приобретают сходство со скелетными. Отличить их можно по наличию прямых переходов от тонкостенных к толстостенным гифам и наличию перегородок. У гименохетовых грибов наблюдаются толстостенные гифы с пигментированными стенками и регулярными перегородками, отделяющими сравнительно короткие клетки. Эти гифы, генеративные по своему происхождению, выполняют в базидиомах механическую функцию и носят название псевдоскелетных (скелетоидных). Такой тип гиф характерен для некоторых видов рода Phellinus.
Скелетные гифы, по определению Корнера, это толстостенные гифы неограниченного роста, развивающиеся из генеративных гиф в зоне роста, как правило, совсем не ветвящиеся или с дихотомически разветвленными концами, без перегородок или с отдельными ложными перегородками. Толстостенность и отсутствие плазменного содержимого указывают на механическую функцию этих гиф в базидиоме. Появляются они не сразу, а после некоторого периода роста и развития. Так как скелетные гифы развиваются в зоне роста и имеют неограниченный рост, их всегда можно видеть в растущем крае.
Связывающие гифы имеют ограниченный рост, развиваются позади зоны роста, сильно ветвятся, как правило, имеют утолщенные стенки, часто извитые; как и скелетные гифы, быстро теряют плазменное содержимое и выполняют в базидиоме механическую функцию. У гименохетовых грибов связывающих гиф нет.
Кроме скелетных и генеративных гиф, в ткани базидиом могут наблюдаться специализированные модификации в виде щетинковидных гиф, траматических простых и звездчатых (многолучевых) щетинок. Щетинковидные гифы квалифицируются как видоизмененные скелетные гифы, так же как и траматические щетинки, отходящие от генеративных гиф. Щетинки, являющиеся специализированными окончаниями скелетных гиф, служат их продолжением и не изменяют «митичность» базидиомы.
Дифференциация гиф в базидиомах не всегда оказывается достаточно четкой, поэтому в сложных случаях некоторые специалисты предлагают использовать более общее деление на генеративные и вегетативные (скелетные и связывающие) гифы (Pouzar, 1966). Э.x. Пармасто (1970) предлагает в аналогичных ситуациях деление на мономитический и гетеромитический типы гифальных систем.
Если базидиомы состоят только из генеративных гиф, тип гифальной системы квалифицируется как мономитический.
На ранних этапах развития базидиом генеративные гифы тонкостенные, но позднее их стенки могут утолщаться. Как правило, это утолщение наблюдается в первую очередь в подстилке или ткани шляпки у нерезупинатных видов, субгимениальные гифы обычно остаются тонкостенными. Большая часть гименохетовых грибов характеризуется мономитическим типом гифальной системы.
Часть видов рода Phellinus имеет настоящую димитическую систему с дифференциацией гиф на генеративные и скелетные. Генеративные гифы гименохетовых грибов бесцветные или слабо пигментированные, в димитических базидиомах остаются тонкостенными и быстро замещается скелетными, так что в относительно зрелых спорофорах их можно обнаружить в растущем крае только при специальном поиске. Скелетные гифы толстостенные, обычно прямые и почти не ветвящиеся, очень редко с ложными перегородками, чаще несептированные, в большинстве случаев параллельно расположенные. При этом свободного пространства между ними практически нет и базидиомы имеют плотную деревянистую консистенцию, позволяющую им противостоять разрушительному действию факторов внешней среды. Димитические виды рода Phellinus обычно ассоциируются с развитием на живых деревьях и способны к многолетнему существованию.
Кроме генеративных и скелетных гиф, в базидиомах гименохетовых грибов встречаются траматические щетинки и щетинковидные гифы. В последнем случае эти структурные элементы выполняют функцию скелетных гиф. Гифальная система, состоящая из генеративных и щетинковидных гиф, носит название щетинкодимитической по классификации Э.x. Пармасто. Такое строение имеют Inonotus niduspici, Phellinus ferrugineofuscus. Траматические щетинки характерны для Ph. ferruginosus и Ph. contiguus. Своеобразное строение отличает Asterodon ferruginosus. В подстилке этого гриба имеются генеративные и толстостенные скелетные гифы, заканчивающиеся звездчатыми щетинками. Таким образом, гифальную систему этого гриба можно рассматривать как димитическую, но если считать звездчатые щетинки самостоятельным структурным элементом, то как щетинкотримитическую (Пармасто, 1970). Настоящая тримитическая гифальная система у гименохетовых грибов не известна.
Тип гифальной системы и связанная с ним консистенция ткани соответствуют образу жизни вида. Однолетние виды могут иметь мономитическую или димитическую гифальную систему, но никогда не бывают тримитическими. В то же время наличие слоистого гименофора, указывающего на многолетний характер базидиомы, одновременно подтверждает наличие более совершенного типа гифальной системы, чему соответствует и консистенция ткани. Виды, развивающиеся в условиях повышенной влажности, имеют, за редким исключением, мономитическую гифальную систему с тонкостенными гифами, в ксерофильных условиях тримитическую, изредка мономитическую с толстостенными гифами. Грибы, развивающиеся на живых деревьях в условиях константной умеренной влажности, большей частью имеют димитическую гифальную систему со скелетными гифами или псевдодимитическую с псевдоскелетными гифами (род Phellinus). Когда развитие базидиом связано с потреблением большого количества влаги, они либо совсем не развивают скелетных гиф (Inonotus), либо механические гифы появляются только после того, как базидиома достигает максимального размера. Базидиомы, растущие на земле, не отличающиеся постоянством в требовании влаги, могут быть мономитическими или димитическими в зависимости от экологии и продолжительности их существования. У мономитических наземных базидиом клетки, как правило, укороченные. Мезофилы на валеже могут быть мономитическими, с толстостенными гифами или димитическими.
Форма базидий имеет таксономическое значение у трутовиков в разграничении крупных групп, однако у гименохетовых грибов стабилизирован булавовидный тип, так что значение имеет только размер базидий, в качестве видового признака. При практическом определении характеристика базидий теряет свое значение из-за трудности наблюдения.
Споры в качестве генеративного элемента играют важную роль в систематике, поскольку отличаются стабильностью формы и размеров и варьируют у одного вида в очень узких пределах. Для гименохетовых грибов характерны тонкостенные, гиалиновые или окрашенные в желтоватые или буроватые тона, цилиндрические, эллипсоидные до почти шаровидных споры. Оболочка гладкая, у некоторых видов толстая двойная. Амилоидность (появление синей или лиловой окраски под действием реактива Мельцера) не наблюдается, изредка споры декстриноидные (окрашивание оболочек в бурый цвет в реактиве Мельцера) и цианофильные.
Из стерильных элементов гимения у гименохетовых грибов регулярно встречаются щетинки. Это особые толстостенные выросты скелетных или генеративных гиф, развивающиеся в субгимении и выступающие между базидиями. Количество их в базидиомах одного вида резко колеблется. В многолетних базидиомах можно наблюдать, что в годичных слоях трубочек количество щетинок, их частота значительно различаются. Особенно это характерно для трубчатых родов. Некоторые виды родов Inonotus и Phellinus совсем утратили способность к образованию щетинок, в других случаях их отсутствие оказывается родовым признаком (роды Coltricia, Phylloporia, Inonotopsis). Вероятно, функция щетинок в гимении связана с механической защитой, но если для рода Hymenochaete такое толкование выглядит весьма правдоподобным, то для защищенного внутри трубочек гимения у представителей других родов надобность в такой защите отпадает. Возможно, у трубчатых грибов из этого семейства щетинки оказываются рудиментарным органом, что подтверждается непостоянством их развития. С точки зрения диагностики имеют значение форма и размеры щетинок. Они могут быть немного крупнее базидий или далеко выходить за пределы гимения (например, гигантские щетинки Phellinus palmicola). По форме различаются прямые шиловидные, конусовидные со вздутым основанием или согнутые, когтевидные щетинки. Иногда в траме наблюдаются щетинки до 100—300 мкм длины, не выступающие в гимении. Они отличаются от щетинковидных гиф ограниченным ростом и известны у некоторых видов рода Phellinus: Ph. contiguus, Ph. ferruginosus. На поверхности шляпок Inonotus cuticuldris встречаются характерные разветвленные или раздвоенные щетинки с крючкообразно загнутыми концами. Их можно наблюдать в волосках опушения.
Иногда в гимении наблюдаются парафизидные гифы и базидиолы.
В старых системах из-за различий в форме гименофора гименохетовые грибы обычно помещали в разные семейства: род Hymenochaete — в Thelephoraceae или Stereaceae, Asterodon — в Hydnaceae или отдельное семейство Asterostromataceae; роды Inonotus, Phellinus, Phylloporia, Onnia, Coltricia — в Poly-poraceae. Патуйяр (Patouillard, 1900) был первым исследователем, признавшим родство гименохетовых грибов независимо от формы гименофора. Все они входят в состав трибы Porohydnes, причем роды Hymenochaete и Hydnochaete включены в подтрибу Pores наряду с родами, имеющими трубчатый гименофор, а род Asterodon со звездчатыми щетинками в ткани отнесен к подтрибе Asterostromes. А.С. Бондарцев и Р. Зингер (Бондарцев, 1953) в своей системе объединяют роды Cyclomyces, Cycloporus, Polystictus s. str., Coltricia, Inonotus, Phellinus в трибе Inonoteae подсемейства Fomitoideae семейства Polyporaceae, добавляя к ним род с белой тканью Leucophellinus.
Донк, последовательно развивая идеи Патуйяра, уже в ранних работах рассматривал грибы с щетинками в гимениальном слое и буроокрашенной тканью, темнеющей в растворе щелочей, в отдельном подсемействе Hymenochaetoideae семейства Aphyllophora-сеае независимо от формы их гименофора. В 1948 г. он описал новое семейство Hymenochaetaceae (Donk, 1948), отличающееся по составу от подсемейства Hymenochaetoideae, описанного в 1933 г. В качестве признаков семейства в диагнозе указаны наличие щетинок в гимении и отсутствие пряжек на гифах. Дополнительным признаком, не включенным в описание семейства, является бурая окраска ткани, темнеющая в растворе КОН. В состав семейства также были включены роды с астерощетинками Asterodon и Asterostroma.
Позднее Донк (Donk, 1964) более тщательно разработал систему семейства, выделив подсемейства Vararioideae, Astero-stromatoideae и Hymenochaetoideae. Последнее включает роды без дихогифид, в виде исключения с астерощетинками, переходящими в гимении в гаплощетинки, в большинстве случаев с гаплощетинками в гимении, иногда с макрощетинками в траме или ткани шляпки. Щетинки могут и отсутствовать. Представители этого подсемейства имеют эвгимений в отличие от катагимения Vararioideae и эвгимения без щетинок Asterostromatoideae. Глеоцистиды отсутствуют, если имеются, то как исключение. Споры гладкие, тонко - или толстостенные, в виде исключения с нежной орнаментацией, неамилоидные, изредка декстриноидные. В подсемейство входят роды с разнообразным строением базидиомы (кортициоидные, клавариоидные, стереоидные, гидноидные, радулоидные до ирпикоидных и полипороидные).
В таком объеме семейство оказалось перегружено родами, близость которых труднодоказуема. Поэтому, очевидно, многие специалисты признают семейство Lachnocladiaceae, выделенное Ридом (Reid, 1965) для грибов с дихогифидами. В более узком объеме семейство Hymenochaetaceae представляется естественным и принято в настоящее время большинством специалистов (Бондарцева, 1983).
Большинство родов, входящих в семейство, имеет однородный видовой состав и невелико по объему. Однако границы между родами не всегда ясные, так же как в некоторых случаях границы между видами. Например, виды рода Inonotus имеют однослойный гименофор, рода Phellinus — слоистый. Однако слоистость у ряда видов неясная, а у тропических видов иногда отсутствует, тогда как «инонотоидный» по структуре базидиомы вид Inonotus weirii имеет часто двуслойный гименофор. Род Phellinus наиболее крупный по объему, представители его одинаково обильны в умеренном поясе и в тропиках, растут на живых стволах, сосуществуя с деревом-хозяином десятки лет, или ведут типично сапротрофный образ жизни. Консистенция ткани у грибов, относимых к этому роду, изменяется от пробково-волокнистой до твердой деревянистой, базидиомы весьма разнообразны по форме. Некоторые виды характеризуются наличием на границе ткани и трубочек тонкого желатинозного слоя — «черной линии». Иногда таких линий несколько. Споры у видов Phellinus с тонкой или толстой, окрашенной или гиалиновой оболочкой, по форме варьируют от аллантоидных, цилиндрических до шаровидных. Тип гифальной системы у большинства видов димитический, но ряд видов имеет регулярно септированные псевдоскелетные гифы. У некоторых тропических представителей (Ph. fastuosus, Ph. calcitratus) даже псевдодимитичность выражена слабо. Все эти данные говорят о гетерогенности рода, однако выявить четкую корреляцию по ряду признаков, что позволило бы выделить более гомогенные таксоны, пока не удается.
Пытаясь разграничить этот сложный таксон, Котлаба (Kotlaba, 1968) предложил виды со щетинковидными гифами, выступающими в виде щетинок в гимении, и бесцветными ацианофильными инамилоидными индекстриноидными тонкостенными спорами выделить в подрод Phellinidium. Кроме того, на основании наличия или отсутствия различных типов щетинок и щетинковидных гиф он отметил в пределах рода 5 групп без таксономической детерминации. В 1980 г. М.А. Бондарцева и С. Эррера предложили систему рода Phellinus из 8 секций и 7 подсекций, используя сочетание 3 эволюционно значимых признаков: окраски спор, типа толстостенных гиф (скелетных или псевдоскелетных) и наличия гимениальных, траматических щетинок и щетинковидных гиф. В конце того же года Риварден (Ryvarden, Johansen, 1980) независимо предложил сходную систему рода, выделив секции на основе наличия или отсутствия гимениальных и траматических щетинок, щетинковидных гиф, поверхностной корки, тонкого желатинозного слоя, а также по форме базидиомы в некоторых случаях. По этим признакам он описал под названием секций 5 групп, но также не придал им таксономического ранга.
Биохимические исследования дали возможность Фиассону выявить в семействе гименохетовых некоторые группы, сходные по содержанию определенных веществ. Оказалось, что базидиомы разных видов различаются по наличию стирилпиронов и гифоломина В (Fiasson, 1983), что дало повод этому специалисту совместно с Ниемеля (Niemela, Fiasson, 1984) предложить новую структуру семейства с относительно большим количеством новых микрородов. Недостатком этих предложений является отсутствие корреляции между морфологическими и биохимическими характеристиками, а значит и возможности экстраполяции выводов на неизученные виды. Поскольку количество исследованных видов пока весьма ограничено, не представляется возможным использовать эти предложения в современной классификации.
Э.x. Пармасто (1985) развивает концепцию микровидов у гименохетовых грибов и описывает много примеров видов-двойников, отличающихся незначительными макро- или микроскопическими особенностями. Разработка этой концепции перспективна и на родовом уровне.
Из всех экологических факторов, определяющих распространение гименохетовых грибов, основным несомненно является субстрат, поскольку большая часть их развивается на древесине. По приуроченности к субстрату можно выделить следующие группы.
1. Виды, растущие на живых стволах древесно-кустарниковых пород, вызывающие стволовые и корневые гнили хозяев: Phylloporia ampelina, Ph. ribis; Onnia leporina, O. triqueter; Inonotus dryadeus, I. dryophilus, I. hispidus, I. iliensis, I. niduspici, I. obliquus, I. pseudohispidus, I. tamaricis, I. weiriv, Phellinus alni, Ph. baumii, Ph. chrysoloma, Ph conchatus, Ph. hartigii, Ph. hippophaecola, Ph. igniarius, Ph. kravtzevii, Ph. linteus, Ph. lundellii, Ph. nigricans, Ph. pinif Ph. tuberculosus, Ph. populicola, Ph. rimosus, Ph. robustus, Ph. torulosus, Ph. tremulae, Ph. vaninii.
2. Виды, растущие на отмирающих и сухостойных стволах, отмерших ветвях живых деревьев, корнях или отмершей древесине в почве, способные развиваться на живой древесной ткани, но предпочитающие отмершую: Onnia tomentosa; Inonotus cuticularis, I. nodulosus, I. polymorphus; Phellinus ferreus, Ph. gilvus.
3. Сапротрофы на древесине: Inonotopsis subiculosa; Coltricia cinnamomea; Inonotus andersonii, I. glomeratus, I. pseudoobliquus, I. radiatus, I. rheades; Phellinus chinensis, Ph. contiguiformis, Ph. contiguus, Ph. ferrugineo-fuscus, Ph. ferruginosus, Ph. laevigatus, Ph. nigrolimitatus, Ph. punctatus, Ph. rhamni, Ph. tricolor, Ph. ussuriensis, Ph. viticola, Ph. xeranticus.
4. Виды, растущие на почве, изредка на сильно разрушенной древесине: Coltricia perennis.
Некоторые из видов Phellinus, паразитирующие на живых стволах, очень быстро прекращают свое развитие после гибели дерева, например Ph. tremulae, другие, напротив, способны расти довольно долго. Род Inonotus образует однолетние базидиомы, как правило, быстро отмирающие после гибели ствола, если находились в жизнеспособном состоянии в этот момент (I. hispidus, I. pseudohispidus, I. dryophilus и др.), хотя некоторые виды, поселяясь на живых деревьях, успешно продолжают развитие на отмерших.
Ряд видов, особенно патогенов живых деревьев, имеет сравнительно узкую специализацию на немногих древесных породах. Это в первую очередь виды рода Phellinus. Ph. igniarius — комплекс представляет собой наиболее яркий пример такой специализации. От исходного вида в разное время были дифференцированы близкие морфологически, но отличающиеся специализацией и имеющие незначительные, но четкие отличия в макроморфологии и отчасти микропризнаках виды: Ph. igniarius, Ph. populicola, Ph. nigricans. Очень близки морфологически и отличаются в основном специализацией Ph. robustus (на дубе) и Ph. hartigii (на пихте).
С состоянием субстрата отчасти связана и продолжительность существования базидиом. В семействе гименохетовых многолетние базидиомы известны для немногих видов, преимущественно из рода Phellinus, причем настоящие многолетние базидиомы со слоистым гименофором развиваются только на живых деревьях. Мицелий (вегетативное тело гриба) всегда многолетний, но базидиомы различаются по продолжительности жизни. В роде Hymenochaete известны виды с 2—3-летним существованием базидиом, все представители родов Asterodon, Phylloporia, Inonotopsis, Inonotus, Onnia, Coltricia однолетние. Базидиомы Coltricia в умеренном климате способны зимовать в виде исключения. По-настоящему многолетние базидиомы известны только в роде Phellinus, однако продолжительность их жизни определяется не только состоянием субстрата, но и климатическими условиями: большая часть тропических видов характеризуется 2—3-летним сроком жизни базидиом, причем слоистость гименофора у многолетних видов выражена слабо. Хорошо заметные слои наблюдаются преимущественно у видов, обитающих на живых деревьях.
По отношению к влажности у дереворазрушающих грибов можно выделить группы гигро-, мезо - и ксерофилов. Большая часть гименохетовых грибов относится к группе мезофилов. Количество ксерофилов ничтожно мало. Наиболее яркий представитель этой экологической группы — Coltricia. Гигрофилы в семействе гименохетовых также немногочисленны, примером может служить Asterodon ferruginosus.
Важнейший биологический процесс споруляции также коррелирует в определенной мере с отношением гриба к температуре воздуха и влажности. У гименохетовых грибов, как правило, 2 периода споруляции: основной в конце лета—осенью и второй период — ранней весной—в начале лета. В умеренном климате у большинства изученных видов споруляция начинается при температуре воздуха 2—3 °С и прекращается, когда начинаются кратковременные ночные заморозки. При 5—10 °С у большинства видов споруляция идет активно. Весенняя споруляция начинается в первых числах апреля и заканчивается в мае или середине июня, осенняя продолжается с конца июля до конца ноября. Изменение влажности воздуха более сказывается на грибах, растущих на мертвой древесине. Виды, развивающие базидиомы на живых деревьях, при снижении влажности воздуха споруляцию не прекращают.
В условиях умеренного климата зимний перерыв в споруляции легко объясняется резким понижением температуры и прекращением вегетации даже у многолетних базидиом; летний перерыв может быть объяснен либо понижением влажности воздуха, либо тем, что в это время происходит интенсивный рост гименофора и базидий не образуются. По данным Э.x. Пармасто, споруляция продолжается у Inonotus radiatus 2—6 нед, у Coltricia perennis — 6 нед, у многолетних видов (в нед): Phellinus igniarius — 2.5—5.5, Ph. tremulae—1.5—5.5, Ph. laevigatus — 4.5—6, Ph. nigrolimitatus — 2.5—3, Ph. pini — 0.5—4, Ph. chryso-loma— 1.5—4, Ph. ferrugineo-fuscus— 1.5—3 в зависимости от погоды.
Поскольку многие гименохетовые грибы связаны с патогенезом живых деревьев, круг пород-хозяев, поражаемых одним видом, как правило, значительно меньше, чем количество пород, поражаемых сапротрофами. Однако далеко не всегда можно встретить соответствующие виды на породах, которые они способны поражать. В каждой местности имеется круг пород, предпочитаемых данным грибом, причем в разных географических районах эти породы различны для одного вида гриба. Наблюдения показывают, что границы распространения древесно-кустарниковых пород не совпадают с таковыми дереворазрушающих грибов, к которым относятся и гименохетовые. Причины такого несоответствия заключаются в том, что в пределах своего ареала гриб переходит с породы на породу. В различных климатических условиях восприимчивость пород-хозяев к поражению грибами неодинакова. Это связано, очевидно, с длительностью вегетационного периода и жизнестойкостью породы-хозяина в данных условиях. Видовой состав грибов в центре распространения древесной породы богаче, чем на границе ареала.
На родовом уровне гименохетовые грибы могут рассматриваться как космополиты, так как встречаются на всех континентах. Узкоэндемичных видов среди них, по-видимому, нет, есть виды, распространение которых недостаточно изучено. Род Coltricia более связан с тропиками, род Onnia — с умеренной зоной. Остальные роды более или менее равномерно представлены в разных географических районах. Из тропических видов большую часть составляют пантропические. В умеренной зоне некоторое количество видов более связано с горными лесами (Phellinus nigrolimitatus, Ph. ferrugineofuscus), хотя они встречаются и на равнинах. Точное выявление ареалов гименохетовых грибов — дело будущего, так как пересмотр видовых концепций ряда распространенных видов сильно изменяет представление об их ареалах.
Все представители семейства Hymenochaetaceae вызывают белую гниль, т.е. способны разлагать лигнин и в меньшей степени целлюлозу. У лигнинразрушающих грибов наряду с гидролитическими ферментами имеются окислительно-восстановительные — оксидазы. Много пероксидазы выделяют грибы из рода Phellinus, особенно Ph. igniarius. Грибная пероксидаза в базидиомах не отмечена и выделяется только вегетативным мицелием. По характеру выделения экзоферментов дереворазрушающие грибы различаются, что доказывается лабораторными опытами. У гименохетовых грибов в этом плане изучены представители рода Phellinus. Выяснилось, что в культуре оксидазы выделяются первоначально только под центральной частью колонии и лишь позднее выходят за ее пределы. В процессе разложения древесины некоторые виды разлагают первоначально лигнин и гемицеллюлозы и только на поздних стадиях развития целлюлозу. В результате древесина приобретает вид сети с ячейками, заполненными белыми хлопьями неразложившейся целлюлозы. Такая гниль называется пестрой или ямчатой. В конечной стадии гниения древесина становится мягкой, волокнистой, часто расслаивается параллельно годичным слоям, теряет в массе, но ее объем не уменьшается. Такой тип гниения характерен для Ph. pini и Ph. chrysoloma. Большая часть гименохетовых грибов вызывает типичную белую гниль.
Данные о биохимических особенностях гименохетовых грибов пока носят разрозненный характер. Споры некоторых видов цианофильны и декстриноидны, амилоидность микроструктур в семействе не отмечена.
Попытка изучения элементарного состава гименохетовых грибов показала, что Ph. robustus (возбудитель белой гнили) и Ph. pini (возбудитель пестрой гнили) образуют подгруппу в семействе. Полипороидный гриб Melanoporia quercina располагается в центре точек семейства Hymenochaetaceae, так же как и Osmoporus odoratus. Желтые стирилпироны характерны для гименохетовых грибов из родов Hymenochaete, Inonotus, Phellinus. Так называемая ксантохроидная реакция, заключающаяся в появлении бурой окраски мицелия или базидиомы под влиянием щелочей, имеет самостоятельное значение в систематике группы. Пигмент, вызывающий эту реакцию, относится к группе фенольных полимеров (Пармасто, 1979). Эта реакция наряду с цианофильностью и декстриноидностью микроструктур имеет диагностическое значение.
Хозяйственное значение гименохетовых грибов определяется прежде всего их развитием на живых деревьях. Количество патогенов древесно-кустарниковых пород в этом семействе больше, чем в других семействах дереворазрушающих грибов. Выше были перечислены виды, вызывающие стволовые и корневые гнили древесно-кустарниковых пород. Хозяйственное значение каждого вида различается в разных географических районах, однако есть виды, которые повсеместно приносят большой ущерб. Во всех районах произрастания осины деревья, начиная с 30—40-летнего возраста, а иногда и более молодые, поражаются ложным осиновым трутовиком Phellinus tremulae. На березах почти повсеместно распространены скошенный трутовик Inonotus obliquus — возбудитель чаги и ложный трутовик Phellinus igniarius. В южных районах нашей страны — на черноморском побережье Кавказа и Крыма, в Средней Азии — широко распространен в садах и парках Inonotus hispidus, поражающий деревья платана, шелковицы и некоторых других пород. В тугайных лесах Средней Азии большое хозяйственное значение имеют I. pseudohispidus на тополях и I. tamaricis на тамариске. В Сибири большие пространства заражены трутовиками Гартига (Phellinus hartigii) и Вейра (Inonotus weirii). Плодовые насаждения страдают от Phellinus tuberculosus (косточковые и семечковые породы) и Phylloporia ribis (ягодные кустарники). В европейской части страны хвойные породы в лесах поражены сосновой губкой (Phellinus pini). По существу почти все виды, способные развиваться на живых деревьях, имеют хозяйственное значение, так как широко распространены если не повсеместно, то во многих районах.
Меры борьбы с гименохетовыми грибами носят в основном профилактический характер, так как реально совершившееся заражение становится очевидным тогда, когда единственным способом борьбы с грибом остается рубка. Правильная лесоэксплуатация, рубки ухода, а в необходимых случаях санитарные, соблюдение культуры лесоразведения способствуют созданию здоровых насаждений. В некоторых случаях полезны работы по селекции на устойчивость к поражению стволовыми гнилями некоторых древесных пород, имеющих большое хозяйственное значение. Например, известно что в некоторых районах, особенно горных, осина не поражается грибом Phellinus tremulae. Устойчивыми оказываются преимущественно зеленокорые формы. Поскольку эта быстрорастущая порода находит разнообразное применение в народном хозяйстве, выведение устойчивых к поражению форм было бы вполне оправдано. В садах и парках, на ограниченной территории, при поражении особо ценных деревьев применяется сбор базидиом со стволов и пломбирование дупел. Уменьшение рекреации в ценных лесных массивах также способствует их сохранению.
Мощный ферментативный комплекс гименохетовых грибов в принципе может быть использован при биологическом способе получения целлюлозы, однако на практике более перспективны быстрорастущие в культуре и более активнодействующие виды истинных сапротрофов из других семейств трутовиков. Паразитические свойства гименохетовых грибов служат причиной их недостаточной активности при разложении древесины в лабораторных условиях.